1872年,在意大利一家廚房裡發生了神經科學中的一次重大進展。帕維亞大學年輕的醫學研究生卡米洛·高爾基(CamilloGolgi)由於對大腦的強烈興趣而建立了一個簡易實驗室。困擾高爾基的問題是關於物質腦的本質:腦是由什麼組成的。那時,儘管可將大腦切成碎片,但在顯微鏡下只能觀察到一堆均質的蒼白色漿狀物。除非能夠鑒定出腦的基本構件,否則不可能發現它是如何工作的。然而,隨著故事的發展,有一天,高爾基偶然地將一塊腦塊放入盛有硝酸銀溶液的碟子中,並在其中浸泡了幾個星期。結果,高爾基發現了一個極其重要的反應。當他取出腦塊時,變化已經發生了。在顯微鏡下出現了一種複雜的圖案:在網狀的纏結中懸浮著黑色的斑點。我們現在知道,一旦將腦組織放入硝酸銀中三個小時或更長,就有可能顯現出腦組織最基本的組分——特殊類型的細胞,這種細胞稱為神經元。
高爾基的發現中更令人不可思議的是,通過一種目前仍然無人完全知曉的變化莫測的過程,染色只隨機地標記出十分之一到百分之一的細胞,因此表現為蒼白的黃褐色背景上的黑點。如果每一個神經元都被染色,那麼精細、複雜的細胞輪廓,將被其他細胞相互重疊的部分所掩蓋,腦組織在顯微鏡下的整個視野裡將變成幾乎均勻的黑色。由於僅有百分之一到百分之十的細胞與高爾基染料起反應,這些細胞即因反差明顯而凸現出來。
神經元到底像什麼呢?在所有的神經元中,都有一個直徑約50微米粗短的團狀部分,稱為腦體(soma,來源於希臘語「身體」)。實際上,胞體的形狀並不像團狀一樣模稜兩可和無定形,而總是屬於以下幾種特徵性形狀之一,如圓形、卵圓形、三角形甚或梭形(形如老式的紡錘)。胞體包含了神經元生存所必需的所有細胞器。從這點上講,神經元的胞體與其他任何細胞並無差別。然而,如果將神經元與其他細胞相比,一旦注意到胞體以外的部分,你就會發現一個巨大的差異:與其他細胞不同,神經元除了胞體外還有其他部分。
纖細的分支從神經元胞體中伸出,幾乎就像某種微小的樹。事實上,這些部分被稱為樹突(dendrites,來自希臘語,意為「樹」)。一個神經元的樹突在形態上千姿百態,在密度上千差萬別,它們或者從神經元的四周長出使其呈星形,或從胞體的一端或兩端伸出。根據樹突分支的程度,神經元在總體外觀上差異懸殊:在腦中,神經元至少有50種基本形狀。
神經元不僅有這些小分支,而且絕大部分還有一個從胞體上伸展出來的長而細的纖維,稱為軸突,它要比神經元其餘部分長許多倍。一個細胞的直徑通常約20至100微米,但在一種極端的情況中可長達一米,如沿著人脊髓下行的神經纖維。
只要看一眼神經元,你就很容易區分出這兩個特徵來。由於軸突遠細於相對粗短的、分叉的樹突,甚至在顯微鏡下也極難看到它們。樹突就像真的樹上的分枝,其末梢逐漸變細,軸突則不然。這就使得神經元在總體外觀上表現為一個團狀的中心區、一根蜿蜒而行的細長的纖維,以及伸出的相對粗短的微技。如此怪異的東西怎麼可以成為我們的個性、希望和恐懼的構件呢?
既然胞體中含有與所有其他細胞相似的整套內部裝置,很容易推想,至少它的某些功能是為了確保細胞存活,並製造出適當的化學物質。然而,鑒於軸突和樹突的存在與神經元的特殊功能緊密相關,因此它們的作用並不那麼一目瞭然。此外,軸突和樹突間如此清晰的形狀上的差異,提示它們扮演著迥然不同的角色。
在第二章中我們曾看到,腦電圖在反映腦狀態的變化上十分有效,又相當敏感,那時我們首次接觸神經元能產生電的想法。樹突充當這些信號的接收區,就像某個巨大的碼頭一樣,接納各種船隻載入的貨物。正如貨物可以從碼頭上卸下,並沿著匯聚於某個中心工廠的路線運送,這些分散的信號沿著彙集於胞體的樹突傳導,如果信號足夠強,樹突將會產生一個新的電信號,或者沿用剛才的類比,將會生產出一個新的產品。這時軸突開始起作用:它們將這個新的電信號從胞體傳送到回路中的下一個目標神經元,就好比工廠的產品輸送到某個遠處的目的地。在本章中,我們將考察一下神經元是如何發出和接收電信號的。我們也將看到,特殊的化學物質在神經元通訊中是如何發揮其重要作用的,而藥物又如何影響兩個神經元間的這種通訊。
在神經科學研究中,這種從單個神經元出發的方法被稱為自下而上法。其策略是從底部,即最基本的組分神經元起步,然後去瞭解兩個神經元間的通訊最終如何構成一個複雜的實施功能的整體。與之相反的是自上而下法,其基本框架是從頂部的宏觀系統出發,這可以是一個腦區(參閱第一章),或一種功能(參閱第二章),通過向下探索,並將其分解成更小的子系統,來瞭解其如何與腦的運作相適配。按其應用這些研究方法的多少不同,神經科學家通常被分成兩類。從某種意義上講,在前兩章中我們一直在運用自上而下的策略,因此已瞭解其長處和局限性。在本章中,我們轉至基於單個神經元的還原論的方法。
路易吉·伽伐尼(Luigi Galvani)最先證明自脊髓伸出的神經能產生電。在一次雷雨時,他將青蛙的腿放在一塊金屬板上。令人驚異的是,蛙腿隨著雷鳴電閃而抽搐。於是伽伐尼錯誤地推論所有的電都處於活組織中。他想像肌肉是電的容器,神經則是電的導體。相反,19世紀物理學的先驅法拉第則認識到電是一個很基本的現象。他從用非生物材料所做的實驗中得出結論:「無論其來源,自然界中的電都是相同的」。神經確實是電的一個源泉,但並非其專有。
從字面上講,電流是電荷的流動。在腦中,這樣的流動由下列四類最常見的離子(原子缺少一個電子或擁有一個額外電子)中的任一種運動而產生:鈉、鉀、氯或鈣。這些離子分佈於神經元內(鉀)或外(鈉、鈣、氯)。但它們不能輕易地隨機進出,毋寧說,所有四種離子因細胞膜這個壁壘而被停留在某個位置上。細胞膜不僅是單純的一堵牆,而是由中間夾有脂肪的兩層構成,就像一個奶油三明治。由於離子不能透過神經元膜中間與之不相容的油層,因而沒有離子能夠自由地進出神經元。
結果,離於在神經元內外積聚。細胞內還有其他帶有負電荷的蛋白質。當把離子和蛋白質一起考慮時,神經元膜任一側的淨電荷分佈是不平衡的;正電荷和負電荷的數量是不等的。對於神經元外而言,神經元內偏負。這樣就產生一個電位差,即電壓,用負值來表示,通常約-70毫伏或-80毫伏。
然而,假如這些離子一直停留在一個位置上而不能流動,從而不能產生真正的電流,那麼,保持的電位差也就沒有什麼意義。設想一下,如果一個水閘的一邊蓄積了大量的水,而你卻不能用它。對於細胞來說,要想產生電信號,電荷就必須流動,就像水必須從水閘中釋放出來一樣。為了電荷的流動,離子必須短暫地進出神經元。但是離子怎麼可能穿越那個不通透膜中間的脂肪層呢?
膜壁壘畢竟是能夠被突破的。由大分子蛋白質組成的各種特殊結構跨越膜的兩層:它們充當了特定離子從一個水相非脂肪區(神經元外)進入另一區(細胞內)的橋樑。然而,由於這些蛋白質通路緊緊地插入穿過膜中間層,它更像一個隧道。在常規神經科學用語中,實際上稱之為通道。
對於一個發送電信號的神經元,帶正電的鈉離子能短暫地進入細胞,暫時使細胞內部的電位比外部更正(去極化)。然而,這個電位一旦變正,如+20毫伏,帶正電的鉀離子就離開細胞,暫時使膜電位比正常時還負(超極化)。這樣,當一個神經元以這種方式被激活時,電位差將產生一個短暫的特徵性的變化:一個正脈衝,緊隨一個負的超射。這個瞬變的正-負波通常持續約1∼2毫秒,即所謂的動作電位。動作電位不同於電壓(靜息電位),後者一般在細胞不發送信號時總是存在的(見圖5)。
為什麼首先是鈉通道突然開放?換一種提問法,是什麼觸發了動作電位?畢竟一個完全隨機產生的信號是毫無意義的,就像夜間不意中響起的電話鈴聲,而電話線的那一端卻又無人撥打。讓我們回到樹突這種從胞體中長出的樹枝狀突起上來。樹突充當了來自其他神經元的輸入信號的接受站。假如信號相當強,並且(或者)是持續性的,它們將沿著樹突分支一直被傳遞到胞體,就像家裡的電流沿著絕緣不佳的電纜流動一樣。
如同某個大型鐵路站,在任何時刻都有幾十個、幾百個甚或幾千個信號彙集到中央細胞體上。在任一時刻,這種信號串都會影響細胞產生自身電信號(動作電位)的可能性。當這些輸入信號到達胞體時,它們形成很高的總值,即最終的電壓變化。如果在靶細胞中,這個新的淨電壓差足夠大,它將引起對正向電壓變化敏感的鈉通道的開放,又一次產生了一個新的動作電位,但這一回是在第二個神經元中。
對任一給定的神經元,動作電位的大小總是相同的,典型值約90毫伏。但這樣的一致性帶來了一個問題。假如輸入信號變得更多或更強,而接收的神經元由於受制於只能產生一種信號,則它將如何去傳遞這些差別呢?由於動作電位不能變大,當它所接收的信號變強時,神經元則將產生越來越多的動作電位。當這種情況發生時,我們說該神經元更興奮。一個神經元發生信號強弱的方式是由動作電位產生的頻率變化反映的。較常記錄到的神經元速率約為30到100赫茲,但某些神經元放電能高達每秒500個動作電位(500赫茲)。每秒只產生一到兩個動作電位的神經元被視為慢放電的。
為了同它們各自的靶細胞通訊,腦中絕大部分神經元以這種方式產生動作電位。下一個關鍵步驟是動作電位抵達其指定的目的地。就如樹突充當神經元的接收區一樣,更細的單一軸突擔當繼續輸出電信號的輸出通路。電信號(動作電位)傳遞的速率因軸突的直徑及是否為稱為髓鞘的脂肪鞘所絕緣而異。如果髓鞘變性,神經纖維傳遞電信號將不那麼有效,多發性硬化就是這種情況。正常的運動是如此的迅速和自動化,致使幾乎不允許大腦的思維和肌肉的收縮間存在延遲。大腦加工過程和我們運動所具有的輕而易舉的敏捷程度,能夠用神經傳導速度來解釋,它可以高達每小時近352千米!
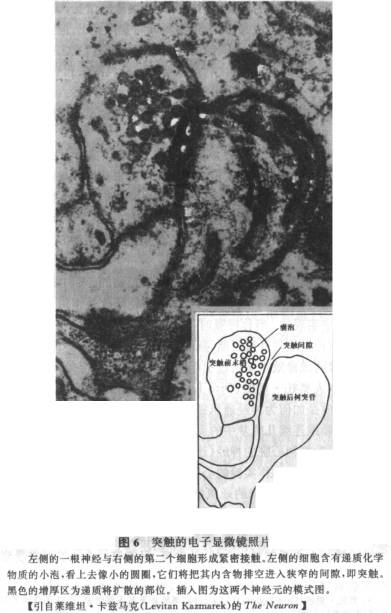
儘管人們已經瞭解神經元中信號是如何產生,又如何沿軸突向下傳遞的,但下一步發生什麼還遠沒有搞清楚。現在我們需要知道的是,為了傳遞信號,神經元實際上是如何同另一個神經元相聯繫的。自從人們能將神經元染色,並加以觀察時起,科學家們就一直思考著這個問題。例如,高爾基認為,所有的神經元是聯結在一起的,有點像一個發網。當時,他遇到了來自西班牙的偉大解剖學家拉蒙·卡赫「(Ramon y Cajal)的激烈反對。這兩個神經科學先驅者在這一長時期的衝突中各執一詞。與高爾基的觀點相反,卡赫確信,在神經元間有一間隙。這個問題直到20世紀50年代才明確地解決,當時出現了一次驚人的突破:發明了電子顯微鏡。
電子顯微鏡所賦予的驚人放大倍數使人們能夠從事對細胞的研究。一台光學顯微鏡採用正常光波和高放大率的透鏡,只能將物體放大到一千五百倍,而一台電子顯微鏡則能放大幾萬倍。科學家用能阻擋電子的特殊物質包埋腦切片,這種物質然後為神經元的不同部分以不同程度吸收。在電子顯微鏡中,一束電子穿過腦組織投射到膠片上。細胞某部分的電子越緻密,其在膠片上就越黑。在電子顯微鏡照片中,神經元失去了它們精細的花朵似的外觀,而是體現了現代藝術一種單色調形式的抽像美。對由清晰的黑線和圓形形成的明確圖案,未經訓練的眼睛分不出它們是軸突、樹突還是胞體。但對於神經解剖學家,神經元的不同部分,就連其內部的細胞器最終也是可以辨認的。
一旦科學家們能以這樣的精確程度去窺視大腦時,神經元的一個秘密就大白於天下了。最終結論是,卡赫是正確的:在兩個神經元間確實存在間隙,即突觸。在腦中,通過細胞不同部位的各種方式的相對排列,神經元相互間形成突觸接觸:樹突間、軸突間都能形成突觸;軸突也能直接與靶細胞胞體形成突觸。最常見的突觸形式是,細胞的離心部分軸突抵達其終點軸突終未,並與目標細胞的粗短分枝部樹突形成突觸(見圖6)。
突觸的概念立即引發了一個問題。試想一下,一個電脈衝信號以每小時約352千米的速度到達軸突終端突觸的情況。確實,這個軸突終端(軸突末梢)被興奮,電位在剎那間變得更正。但這個興奮波何去何從呢?當它為一個間隙(突觸)短暫阻擋後,它怎麼能用作為另一個神經元的信號呢?這有點像駕駛一輛汽車來到一條河邊,一個理想但過分奢侈的對策是放棄汽車,尋找更合適的旅行工具:一艘船。我們需要一種將電信號轉譯成另一種能通過突觸的信號的方法。
自19世紀以來,人們想像,化學物質可能以某種方式參與神經元通訊。南美州印第安人用於狩獵的一種毒藥的效果使法國人克勞·巴納爾(Claud Barnard)入迷。獵人將他們的箭頭浸在一種叫箭毒的物質中。當箭射入獵物身體中後,它不是立即殺死它,而是將其麻痺。巴納爾認為,這種致命的毒素是通過某種方式干擾了體內的神經而起作用的。
直到20世紀初,人們才證明巴納爾是正確的。人們發現,箭毒的作用是通過阻斷一種自然產生的,由神經釋放並作用於肌肉的化學物質來實現的。當你呼吸時,隔膜在神經支配下上下運動。顯然,如果該神經的信號被阻斷,隔膜就無法運動,你就不能呼吸。因此,該毒素的致命效果是通過引起呼吸困難而最終導致死亡。1929年,奧地利人奧托·勒維(Otto Loewi)明確證明,這種自然產生的化學物質是如何為神經通訊提供關鍵性聯繫的。這個故事的發生得益於勒維連續幾晚夢中的靈感。他第一步是去重複已知的東西:如果刺激抑制心臟的神經(迷走神經),那麼心跳就會減緩。重要的是,要知道勒維作實驗的心臟和相伴的神經並非在體內,而是被分離後置於一個特殊的通氧的槽中,並浸浴於一種與正常體液相似的溶液裡保持存活。
這個實驗絕妙的一著是,勒維將浸浴原來那個心臟的溶液,轉而浸浴第二個未受刺激的心臟。他發現,儘管第二個心臟並未受到刺激,其跳動也會減慢。對這個發現唯一的解釋似乎是,一定有某種化學物質在第一個心臟受刺激後釋放到溶液中去。這樣,由於同樣化學物質的存在,當用該溶液去浸浴第二個心臟時,其效果與對第一個心臟的效果相同。我們現在知道,這種化學物質是乙□膽鹼,箭毒阻斷的正是它。乙□膽鹼是腦中不同類神經和神經元釋放出來的許多化學物質的原型,在腦信號傳導中充當重要的使者,專業術語稱為遞質。
乙□膽鹼對心臟作用的發現,對於瞭解腦細胞如何在突觸部位進行通訊具有重要意義。一旦我們認識到電刺激能引起神經釋放一種自然發生的化學物質,我們就能更容易地明白當軸突末梢被電信號有效刺激時,腦的一個突觸上可能會發生什麼。一旦動作電位到達軸突末梢時,它立即建立起適宜的條件,使乙□膽鹼釋放至突觸中。
在產生信號的細胞的神經終端的軸突末梢中,乙□膽鹼儲存在許多囊泡中。當動作電位沿著軸突傳播到達這個末梢區時,這個瞬間的電壓變化觸發一些囊淚把內含物(乙□膽鹼)排空至突觸中。到達的電信號越多,排空的囊泡就越多,釋放的乙□膽鹼也就越多。用這種方式,原先的電信號真實地被轉換為化學信號。動作電位的頻率越高,釋放的乙□膽鹼就越多。
乙□膽鹼一旦釋放,它在所有神經元外部的水相鹽溶液(細胞外溶液)中極易擴散,越過突觸就像渡船過河一樣輕而易舉。然而時間尺度上差異巨大;由於這樣的化學物質是些相對小的分子,在幾毫秒內就可越過突觸間隙。但這種化學物質,如乙□膽鹼,僅僅是一個分子,它實際上如何傳遞信息呢?
讓我們回到汽車和船的類比上來。一旦我們過了河,並將在陸地上繼續我們的旅行時,理想的情況是放棄船而再以汽車代之。原來已轉換成化學信號的電信號,現在需要轉換回電脈衝。我們需要瞭解,乙□膽鹼或任一遞質如何導致靶神經元產生電壓的瞬變。
一旦到達突觸的另一邊,每一個遞質分子必然同靶神經元產生某種接觸。遞質像船一樣需要停靠。在靶神經元的外部有特殊的大分子蛋白質稱為受體。它們是為一種特殊化學物質定制的,精確得就如同鑰匙配鎖或手和手套相配一般。
一個受體不是簡單地讓任何化學物質與之適配,它必定是特異匹配的,即分子構象完美地相對應。一旦遞質被鎖入受體並與之結合,一種新的化學物質,即原來兩個分子的復合體便誕生了,它將觸發以後一系列事件的展開。
對於鈉通道或另一種離子通道開放來說,遞質分子和靶細胞上受體蛋白的結合相當於發令槍。在靶細胞中,這些離子中任一種的進出,都將反映在電位差的瞬變上。接下去,這個電位差的瞬變便成為許多電信號中的一個,沿著樹突傳向胞體。
在某種意義上,我們已走完一個輪迴。當這個特殊的電信號到達細胞胞體時,它立即和其他成千上萬個輸入信號一起,導致靶細胞電壓最終的淨變化。如果這個淨變化足夠大,則該胞體附近的鈉通道又將打開,從而在新的靶細胞中形成一個動作電位。這個新的靶細胞本身又將發出一個信號,成為成千上萬個信號中的一個去衝擊再下一個靶細胞。就這樣,電和化學事件不斷重複,生生不息。
在我們的腦中有多達幾千億個神經元。為了感覺一下一千億究竟有多大,亞馬孫雨林提供了一個恰當的類比。亞馬孫雨林方圓7000000平方千米,約有一千億棵樹木。大腦中的神經元數量基本上與之相當。比喻毋需在此打住。如果我們現在考慮神經元間連接的巨大數量,那麼我們可以說,其大約和亞馬孫森林中的樹葉一樣多。要去想像整個腦中化學活動和電活動的激烈狀態,事實上是不可能的,即使在任一時刻,我們的上千億個神經元中僅有10%在活動。
總之,我們的腦為何以這種方式工作,並不是一目瞭然的。畢竟它需要大量的能量去組裝遞質,而這一過程需要一系列複雜的化學反應的參與。進而,只有當遞質完成其工作後迅速地從突觸中被清除掉,這個電-化學-電信號鏈方能正常地運轉。而甚至連這個遞質清除的過程也需要能量,這是因為遞質為細胞重吸收需要耗能,而使遞質分解的神經元外的□也是耗能的。
這種化學信號系統的另一個問題是時間。我們已經知道,小分子將迅速地擴散越過突觸,但整個突觸傳遞過程耗時幾個毫秒。假如神經元融合在一起,且僅通過傳導電脈衝來工作,那麼這會快得多。事實上,存在一些神經元間的接觸,在這些部位,神經元相互融合在一起,毋需化學突觸。令人啼笑皆非的是,至少在這些情況下,高爾基倒是正確的。在這種設計中,毋需使用遞質,電信號通過這些低電阻接觸(縫隙連接)輕而易舉地、迅速地傳導。這種電傳導不僅快捷得多,而且毋需耗能的化學物質。然而,腦中大部分突觸是化學性的。因而儘管從表面上看化學傳遞費時又費能,但它必定具有其極大的裨益。
再考慮一下有多少個輸入能與一個靶細胞形成突觸:多達十萬個。正如我們所看到的,在每一個突觸,遞質釋放量根據到達每一個輸入末梢的動作電位的數量而定。神經元的激活狀態不是固定的,而是能變化至多達十萬個不同狀態。而且,通過多種不同的化學物質對各自特製靶的作用,不同的遞質對最終電壓具有不同的作用。相反,電傳遞將受限於每一個神經元接頭的被動傳導特性。與化學傳遞相比,電傳遞雖快而經濟,但遠遠缺少可變性和多樣性。化學傳遞則賦予腦功能的極其多樣性:不同的化學物質在不同的時間有程度不同的作用。
有時,一種遞質在神經元通訊中能發揮更精細的作用。儘管不能傳送信息,但它能左右靶細胞對一個輸入信息最終作出的反應。這種左右神經元信號的作用稱為神經調製。與現在熟知的突觸傳遞事件相比,神經調製的概念相對比較新,它給化學性信號傳遞增添了更多的能力。在經典的突觸傳遞中,我們考察的是在某個抽像時刻的單個事件。左右靶細胞反應的概念為特定時間框架增添了另一特性:首先一個事件發生,即調製作用,然後另一個,即實際信號,它將被加強或減弱。神經調製是對經典突觸傳遞的補充,就像錄像片與快照一樣。想要通過細胞間簡單的被動電流擴散來使調製作用在某一特定時期介入,幾乎是不可能的。
至少在我看來,正是腦功能這種化學特異性的特徵使那些企圖用計算機模擬腦的人望而卻步。如果我們用高倍電子顯微鏡看一下神經元網絡,它更像某個盛有一大難纏結的麵條的大鍋,其中包埋著神秘的隆起,而非一塊集成線路板。而且,撇開化學上的多樣性不說,大腦中還有精確的連接,而線路板只能是一個蒼白無力的模擬。匯聚到一個細胞上的不同輸入將導致不同化學物質的釋放,而這些化學物質在任一時刻都具有活性。另外,根據這些輸入強度的大小,將釋放不同量的遞質。最後,每個遞質將裝入自身的受體中,而這一受體都以自己特有的方式去影響靶細胞的電壓。這樣,利用遞質分子不同的組合,在每一個環節都為腦中巨大的靈活性和多樣性提供了空間。
這個分子交響曲是計算機中的景況所無法比擬的。首先,也是最明顯的,腦基本上是一個化學系統,甚至連它產生的電也來自化學物質。更顯著的是,除了進出神經元的離子流外,在細胞內部繁忙而封閉的世界中,不停地發生著大量的化學反應。在這些事件中,有一些將決定細胞對未來的信號的反應如何,但這些事件並沒有直接的電的對應物,或者說與計算機沒有簡單的可類比之處。
其次,神經元本身的化學組分是變化著的,因而沒有分離且不變的硬件,這與可編程的軟件形成鮮明對比。進而,大腦這種不停變化的能力導致了其與硅片系統的第三個差異:計算機對同一個指令的反應幾乎總是不變的,不能產生新的反應,雖然它是能「學習」的。
確實,高級機器人裝置似乎能組織和重組它們的線路,去適應一定的輸入,但它們仍遵守一套已程序化的規則——算法。大腦並不一定按算法運轉:例如,什麼是常識的規則?物理學家尼爾斯·玻爾(Niels Bohr)曾訓誡一個學生,「你不是在思考,只是在作邏輯推理」。事實上,根本沒有外部的智力為大腦編程。它是前攝性的,自發地運轉著。當決定去散步時,只是因為它「喜歡這樣」。計算機能做一些大腦能做的事情,但這並不證明這兩個實體用相似的方法工作,或服務於相似的目的。一台無所事事的計算機沒有執行其主要功能,而一個不在幹事的人則有可能正在經歷一次頓悟。
考察神經元間化學通訊產生的另一個見解是,你能意識到為什麼將自上而下的方法和自下而上的方法協調起來是如此的困難,為什麼將單個突觸發生的事件外推到腦的某種功能是如此的困難。腦是由單個神經元組成的,而神經元處於複雜性不斷增加的回路之中。這些連接不像一排手拉手的人,或在兒童遊戲中從立隊的一頭到另一頭傳送一句信息時,最終是完全走了樣。請記住,與任一特定的神經元形成接觸的神經元在一萬個至十萬個之間。反過來,任一特定的神經元將成為神經網絡中下一個細胞的成千上萬個輸入中的一個。假如我們取只有一根火柴頭大小的腦塊,其表面上的連接就能高達十億個。
只考慮大腦的外層——皮層。假如你用每秒一個的速度數其神經元間的連接,它將花去你三千二百萬年!請記住,人類進化僅始於七百萬年前,因而你不得不用四倍於人類進化的時間來數完它。至於皮層中連接的不同組合的數量,它將超過整個宇宙中帶正電的粒子數!
整體腦功能與任一突觸或任一類遞質間沒有簡單的一一對應關係。可用一個粗略而簡化的類比來說明:一支交響曲同一把小號的聲音並無直接的對應關係。同時從自上而下和自下而上的視角來研究大腦的一種方法是考察藥物的作用。我們能瞭解這些藥物怎樣影響行為,同時也能瞭解在單個突觸部位它們怎樣改變化學通訊。畢竟,貌似個性化的不變的精神完全受命於我們的物質大腦,我們的神經元。
在人們用來改變自己感覺的許多藥物中,尼古丁可能是最常用的。尼古丁在吸第一口煙的十秒鐘內到達大腦,我們有可能看到吸煙者的腦電圖中的即時變化,它變得不同步了(見第二章),表明處於不那麼放鬆的狀態。
尼古丁實際上作用於一種受體,即一個在正常情況下為遞質乙□膽鹼保留的停泊區。這種作用可作為藥物起作用方式的一個例子,它模擬一種天然遞質的作用。然而,有兩個理由表明,這種模擬與其說是乙□膽鹼正常作用的仿真,倒不如說是一種拙劣的模仿。首先是受體刺激的量遠遠高於正常情況下乙□膽鹼本身的情況。在腦中,乙□膽鹼受體的重複和過度的刺激,將對腦功能產生長時程作用:當它們受到的刺激遠遠高於正常情況下來自乙□膽鹼的刺激時,受體將變得越來越不敏感。由於靶神經元變得習慣於這些人為的化學物質的高劑量,神經元逐漸變得習以為常,它們在正常量的乙□膽鹼的情況下卻不能正常發揮功能。因而,需要藥物提供超常的刺激。這便是藥物成癮的化學基礎。
其次,乙□膽鹼作用於幾種不同的受體以產生更為均衡的作用,而尼古丁僅作用於一種受體,從而再次導致一邊倒的效應。這種失衡作用也會以更普遍的方式發生在大腦以外:尼古丁使人處於一種戰時狀態,隨時準備戰鬥或飛行,心跳加快,血壓升高。或許,將吸煙者處於戰鬥或飛行狀態反饋至大腦中這一事件本身是令人興奮和愉悅的。但更普遍的是,由於他們的大腦正在發出信號顯示其受體需要更多的刺激,吸煙者因而只是不斷地吸煙。
另一種拙劣模擬天然遞質的藥物是嗎啡(morphine),雖然其模仿的遞質並非是乙□膽鹼。嗎啡由一種特殊的罌粟製成,因為它使人瞌睡和鬆弛,故用睡神摩耳甫斯(Morpheus)來命名。海洛因是嗎啡的一種衍生物,由於經過化學修飾而更容易進入大腦,因此海洛因癮君子們更喜歡這種藥物:它更易進入大腦,使他們更快地「過癮」。海洛因的副作用包括,瞳孔收縮、便秘和咳嗽反射的壓抑。事實上,正因為後兩種作用,它曾經作為一種有效的藥劑來治療腹瀉和咳嗽。
嗎啡和海洛因的一種致命作用是,通過對腦幹中控制呼吸的呼吸中樞的直接抑制來減慢呼吸頻率。呼吸中樞正處在脊髓上方。有時,這種作用能嚴重到使呼吸停止而導致死亡。事實上,呼吸的抑制是海洛因濫用急性致死最常見的原因。
除了上述顯而易見的危險性外,嗎啡在臨床上最有價值的作用是緩解疼痛。它是所知道的最有效的止痛劑,但因其成癮特性,它僅用於疾病晚期或治療極其嚴重或慢性的疼痛。有一種關於疼痛本質的吸引人的見解來源於服用嗎啡的患者:他們常說他們能感覺到疼痛,但疼痛卻不再困擾他們。這個作用能解釋健康無疼痛的人為什麼仍服用海洛因來尋找「愉悅」嗎?或許,在相似方式下,海洛因濫用者不再為日常生活中的擔憂和焦慮而煩惱。服用海洛因這個理由很快地為另一個所取代,只是為了滿足一次需要而已。
讓我們再回到突觸上來。嗎啡的作用方式是模擬我們身體內某種天然產生的遞質。嗎啡在分子結構上與其極其相似,使其很易嵌入到那個為這些特定遞質(稱為腦啡□、內啡□、強啡□,見第二章)特製的受體蛋白質中。這樣,藥物騙過了靶神經元,使其誤認為正在為天然化學信使所激活。我們體內含有這些天然的物質,它們正如我們所看到的乙□膽鹼那樣,也在一定的神經元間傳遞信號。這是二十世紀七十年代的一個重大發現。而且,這種正常的通訊至少有一些在正常狀態下對緩解疼痛是重要的。例如,如果腦啡□的作用為某種藥物(納酪酮)所阻遏,那麼我們的痛感知覺就更糟。同樣,納酪酮至少阻斷了針刺的某些鎮痛效應(見第二章)。因為內啡□介導針刺鎮痛作用,這就可以解釋為什麼這種作用啟動緩慢,為什麼甚至在撤針後這種作用仍維持。或許,扎入穴位的針之所以能發揮作用,部分是由於引起了內啡□和有關化合物的釋放(見圖7)。
在腦中,使用海洛因和內啡□的自然作用之間有什麼區別呢?因為在我們的腦中有一種天然產生的嗎啡/海洛因的類似物,我們根本沒有藥物成癮的危險。阿片□和嗎啡間的差別類似於乙□膽鹼和尼古丁之間的差別。少量的阿片□將在不同時間內在腦的不同部位釋放出來。然而,當一種藥物(如嗎啡或海洛因)服用後,它將立即作用於所有可能的腦區的每一個突觸:它將侵佔正常的受體位點。受體這種激活過度所造成的後果是,使其習慣於較高劑量的化學物質,而對正常量則變得很不敏感。這個下調過程持續進行下去,直到需要越來越多的藥物來產生與之相同的初始效應。藥物成癮再一次成為不可避免的後果。
另一種危險的成癮藥物是可卡因。它同新的強效可卡因相似,後者因經化學修飾而能用嘴吸而不用鼻嗅。可卡因來源於生長在海拔1000到3000米的安第斯山脈上的可可灌木。這個高原上的秘魯居民每年消耗近九百萬千克的這種樹葉,他們咀嚼或舐食這些樹葉是為了感受它們產生的快感。可卡因作用於腦中一種叫去甲腎上腺素的化學遞質。不像尼古丁或嗎啡直接模擬遞質乙□膽鹼或內啡□的作用,可卡因通過另外一種方式起作用。該藥物增加了自然產生的遞質的有效性。在正常情況下去甲腎上腺素在作用結束後從作用位點被清除掉,可卡因阻斷了這個過程:它阻斷了神經元把遞質攝入其內部,使遞質產生不正常的持續性作用。
可卡因是危險的,因為它不僅增加了腦中去甲腎上腺素的有效性,而且還提高了該遞質在體內其他發揮作用的部位中的濃度,這些部位正是神經支配關鍵器官的地方。去甲腎上腺素的作用是把機體引入一種虛假的應激狀態。心跳加快,血壓上升,從而有產生中風的危險。苯異丙氨(脫氧麻黃鹼)在這一方面有相似的作用,它導致去甲腎上腺素及其前體多巴胺過量釋放。此外,通過阻止其被重攝入神經元,苯異丙氨更能增加這些強效遞質的有效性。這樣,在突觸處,多巴胺和去甲腎上腺素起作用的時間比它們應有的長得多。
多巴胺、去甲腎上腺素,甚至乙□膽鹼,都是從原始腦部(腦幹)的神經元簇中釋放出來的,而這些神經元湧泉般地瀰散投射到皮層和鄰近的皮層下結構等更複雜的區域(見第一章)。這些大範圍的化學遞質系統與覺醒水平有關,包括睡眠和清醒。此外,它們能夠左右、調製整個大腦的神經元活動。因此,能改變這些系統的藥物也將能調節覺醒水平,這並不奇怪。例如,服用苯異丙氨者不能保持安靜,他們注意力無法集中,不斷地為外部環境中出現的平淡無奇的東西和事件所分心。在許多方面,服用苯異丙氨者類似於精神分裂症患者,他們總是為外部世界所擺弄,沒有健全的頭腦,無法對正在發生的事情作出恰當的評估。
第三種被濫用的藥物是迷幻藥(3,4-亞甲基二氧異苯異丙氨,MDMA),它作用於第四個遞質系統(5-HT,即5-羥色胺),該系統也從腦幹向上、向外投射。這種藥常被視為致幻劑,因為它不但能帶來超越一切的狂喜,而且能給人一種靈魂脫殼的感覺。這種藥使5-羥色胺過度釋放。腦中5-羥色胺的氾濫成災會在代謝和體溫調節上產生嚴重的後果。除了產生欣快感外,這種藥也會導致活動亢進。這種不間斷的重複運動正是瘋狂勁舞者的特徵,而這些人往往服用迷幻劑。事實上,在大鼠上可看到相似的效應。把正常大鼠置於箱中,它們不斷地在箱中探查,表現出各種不同的動作,如流眼淚、打噴嚏、行走和洗刷。然而,當大鼠服用了迷幻藥後,它將一遍又一遍地重複同一個動作,與其正常行為相悖。這種重複的運動使人想起受藥物影響者的重複的舞蹈動作,令人毛骨悚然。
現在仍然不知道迷幻藥的作用是否是由於5-羥色胺大量的釋放,或者是因為由此導致的遞質耗竭所致。不管怎樣,需要認真考慮的是,現在有證據顯示,長時間重複使用迷幻藥會導致大鼠神經元核團(中縫核)的死亡。這些神經元從腦幹向上和向外發出噴泉狀的軸突,以瀰散的方式投射到腦的高級區域。它們與一系列極其基本功能的調節有關,包括睡眠。
這些釋放5-羥色胺的噴泉狀神經元也是很多抗抑鬱劑的作用位點。許多抗抑鬱劑是通過增加5-羥色膠的有效性來發揮作用的,但其機制不同,因而其作用方式不會導致神經元的死亡。目前最流行的抗抑鬱劑百憂解(Prozac),就是以這種方式工作的。然而,如果淨增的5-羥色胺有效性伴有「愉悅」,那麼短時服用增加5-羥色胺有效性的藥(如迷幻藥)必將有相似的效果。但與抗抑鬱劑不同,如果重複使用迷幻藥會導致神經末梢死亡,從而使5-羥色胺永久耗竭,因此可以預期,持續使用迷幻藥會有抑鬱的副作用。實際上,有一些資料提示,長時間使用迷幻藥能造成抑鬱和自殺。
我們能夠明白,藥物之所以以多種方法影響大腦,是因為它們作用於大腦中多種化學物質,而且又干預突觸傳遞的不同階段。尼古丁和嗎啡通過作用於某種天然產生的化合物的受體來模擬該物質,而可卡因則擴散過突觸前增加另一種物質的有效性。迷幻藥又不同,事實上它耗竭了腦中另外一種比學遞質。由於我們腦中有如此之多的比學遞質,因此藥物作用方式不但高度多樣化,而且存在高度專一性的靶體。在某種程度上,我們知道藥物能做什麼,但實際上我們並不知道其長時間內的全部作用或對身體其他部位的副作用。
或許,最引人入勝的問題是,已知的分子/細胞水平上的變化和我們真正感覺方式上的變化間存在的聯繫。為什麼由嗎啡引起的內啡□受體的過度刺激實際上會導致欣快的主觀感受和對疼痛的漠視?抗抑鬱劑在突觸水平上的作用(如增加5-羥色胺的有效性)如何能導致抑鬱的緩解?如果我們記得抗抑鬱劑只在幾小時內在生物化學上是有效的,而卻有幾十天的治療效果的話,這個謎就特別令人費解。很顯然,在一種分子和一種特定情緒之間,並不存在簡單的——對應關係。
就此來看,對藥物作用的考察清楚地顯示,突觸所發生的專一明確的事件,以及這些突觸事件最終如何形成一種情緒間聯繫的本質仍然令人難以捉摸。神經科學面臨的一個最重大的挑戰是,如何能用突觸傳遞和化學調製這種自下而上的方法的構件來說明自上而下的現象,如愉快。想像我們獨特的意識如何真正地為腦中的液體和電火花組成的雞尾酒所完全控制,這確實引人入勝。以下兩章將分別介紹個體獨特的大腦是如何組裝起來,又如何表達這兩個問題。
|
|
